
by Marek Ples
|
| Arthur Schopenhauer |
| philosopher (1832) |
|
| Marek Ples |
| hobbyist philosopher (2022) |
In the early days of my first childhood interest in biology, besides paleontology, I was particularly drawn to entomology and related fields (e.g. carcinology, a science about crustaceans). I spent countless hours exploring the world of insects, marveling at their intricate anatomy and fascinating behaviors. As I grew older, I ultimately pursued a different field as my main career path, such as biotechnology or medical engineering. However, my passion for entomology has endured throughout my life, and I continue to study and appreciate the complex world of insects and other invertebrates. Even though my professional focus has shifted, entomology will always remain one of my personal favorites.
Over 1.3 million insect species (without other anthropods) have been described, more than two-thirds of all known species. Some insect species date back to around 400 million years ago. They have many kinds of interactions with humans and other forms of life on Earth.
| Firefly Phausis splendidula syn. Lamprohiza splendidula (ins.) | |
| The Madagascar hissing cockroach Gromphadorhina portentosa (ins.) | |
| The death's head cockroach Blaberus craniifer (ins.) | |
| The common rough woodlouse Porcellio scaber (cru.) | |
| Water flea Daphnia pulex (cru.) | |
| Antlion Euroleon nostras (ins.) | |
| More | |
ins. - insect; cru. - crustacean |
Fireflies have something ethereal about them - they move silently, flickering like lanterns in the darkness. Fireflies have captivated human interest throughout history, symbolizing diverse conditions across different cultures. According to a Japanese legend, lightning bugs are believed to be the souls of the deceased, with some versions of the tale suggesting that they represent the spirits of fallen warriors. The Japanese word for a firefly is hotaru (ホタル). While fireflies may not hold a positive reputation in certain cultures, they are highly regarded in Japanese society. In fact, they have been used as a metaphor for passionate love in poetry since the 8th-century anthology known as Man'yōshū (万葉集) or Collection of Ten Thousand Leaves which is the oldest extant collection of Japanese waka (和歌, classical japanese poetry), compiled sometime after AD 759 during the Nara period. In Japan, they are particularly revered for their aesthetic beauty, leading to the creation of dedicated parks where their mesmerizing glow can be admired.
The Lampyridae family consists of more than 2,000 light-emitting beetle species known as fireflies, lightning bugs, or glowworms. These insects use their bioluminescence, primarily during twilight, to attract potential mates. Initially, light production in Lampyridae likely served as a warning signal in larvae, indicating their inedibility, but it later evolved into a mating signal in adult individuals. Female fireflies of the Photuris genus have developed the ability to mimic flash patterns, enabling them to prey upon males from the Photinus species.
Fireflies are predominantly found in temperate and tropical regions, often inhabiting marshes or wet, wooded areas that offer ample food sources for their larvae. While all fireflies emit light during their larval stage, only specific species continue to produce light as adults, and the location of the light organ varies among species and between sexes.
In the natural environment of my country (Poland), three species of beetles from the Lampyridae family can be found:
During one of my field trips, I encountered a firefly, Phausis splendidula, so I would like to share a few words about this charming insect. In the case of firefly from this species, both males and females have the ability to emit light [1]. This is therefore a significant difference compared to the common glow-worm, where only females exhibit bioluminescence.
The insect is characterized by a very noticeable sexual dimorphism. Males have front pair of wings hardened into brown colored wing-cases (elytra) and a slightly lighter thorax equipped with a transparent plate divided into two parts (Fig.1A). On the other hand, females are usually even lighter in color, wingless and resemble larvae in appearance (Fig.1B). They are also significantly larger than males, reaching lengths of up to 2.5 cm (~1.0 in).
Fig.1 - Phausis splendidula, A - male, B - female
Light production in fireflies is due to the chemical process of bioluminescence. This occurs in specialized light-emitting organs, usually on a firefly's lower abdomen. The enzyme luciferase acts on luciferin, in the presence of magnesium ions, ATP, and oxygen to produce light. Oxygen is supplied via an abdominal trachea or breathing tube. The light organ in males is located on the underside of the abdomen, in the form of two strip-like structures (Fig.2).
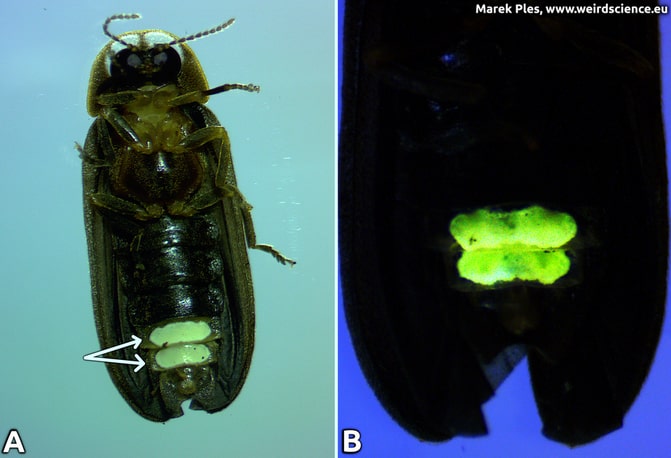
Fig.2 - Light organ of the Phausis splendidula, male; A - localization, B - bioluminescence
Females also have light organs, but in addition to the strip-like structures, they also have glowing spots on the sides of the abdomen (Fig.3).

Fig.3 - Bioluminescence of Phausis splendidula, female
The adult stage or imago of the described firefly lives for a short period, usually up to a few weeks. The most important and essentially sole task during this time is to find a mate for reproduction, and since these insects rely heavily on vision for this purpose, it is not surprising that they possess enormous - relative to body size - compound eyes (Fig.4) [2].

Fig.4 - Compound eyes of Phausis splendidula
While studying the phenomenon of light production in living organisms, in addition to fireflies, I also paid attention to bioluminescent fungus Panellus stipticus, marine dinoflagellates Pyrocystis lunula, and bacteria Aliivibrio fischeri.
Bioluminescence is actually a unique, naturally occurring form of chemiluminescence. We can also conduct this type of chemical reactions in our laboratories (Fig.5).

Fig.5 - Chemiluminescence; TCPO oxidized in presence of perylene (left) and rubrene (right)
Gromphadorhina portentosa is one of the largest species of cockroach, reaching 5.0-7.5 cm (2-3 in) at maturity. They are native to the island of Madagascar, which is off the African mainland. They are commonly living in rotting logs. It is one of some 20 known species of large hissing roaches from Madagascar, many of which are kept as pets, and often confused with one another by pet dealers; in particular, G. portentosa is commonly confused with Gromphadorhina oblongonota and Gromphadorhina picea.
These cockroaches exhibit a shiny brown coloration and have an oval-shaped body. They lack wings and possess a single pair of antennae. The males display large horns, which contribute to their distinctive and impressive appearance (Fig.6).

Fig.6 - Male Gromphadorhina portentosa
To observe the ventral surface of the cockroach's body, it is necessary to immobilize it. The insect should be flipped upside down with its legs, and then covered with a properly sized Petri dish, preferably made of glass. This way, the insect will not be able to flip over, and instinctively it will rest legs against the surface of the glass, allowing for convenient observation of the details of its external structure (Fig. 7).

Fig.7 - External anatomy of Gromphadorhina portentosa, view of the ventral surface of the body; a - antenna, b - compound eyes, c - mouthparts, d - legs, e - sternite, f - abdominal appendages
The most reliable method for determining the sex of a Madagascar hissing cockroach is to examine the last section of their abdomen. If you look at the underside of their abdomen (Fig.8), you will see that their abdomen is made of several sections. For male hissing roaches, the last section ends in a tapered point and is very narrow (<0.6 cm or ~0.25 in crosswise). In contrast, the last section of the female hissing roaches is much larger and round (~1.3 cm or ~0.5 in crosswise).

Fig.8 - Difference in the abdomen structure of the Madagascar hissing cockroach; A - male, B - female; the shape of the last sternites is highlighted with a white outline
The life cycle of the Madagascar hissing cockroach is relatively long and differs from that of most other cockroaches. Females exhibit a reproductive strategy known as ovoviviparity, meaning they give birth to live offspring. The female carries the eggs and newborn nymphs for approximately 60 days until they emerge as first instar nymphs. A single female can produce a considerable number of nymphs, up to 60. This insect follows a mode of development called hemimetaboly or incomplete metamorphosis: egg → nymph → imago (adult). The nymphs go through six molts before reaching maturity, which takes around seven months. Both the nymphs and adults lack wings and can survive for 2 to 5 years.
The Madagascar hissing cockroach derives its common name from its notable ability to produce a distinct hissing sound (Vid.1). This serves as its primary defense mechanism against potential predators. The sound is generated when the insect forcefully expels air through specialized respiratory spiracles, primarily located on the fourth segment of its body (abdomen). While spiracles can be found on various segments of the abdomen, it is primarily from the fourth segment that the hissing sound originates [3]. It is worth noting that the Madagascar hissing cockroach is the only member of its group of cockroaches that is capable of producing audible sounds. This method of sound production differs from that of crickets and other insects, which typically create noises by rubbing body parts together (stridulation), often involving legs.
Vid.1 - Sounds produced by the Madagascar hissing cockroach
Due to its beauty and behavior, the Madagascar hissing cockroach is an excellent insect to study. Additionally, its breeding process is relatively straightforward. Moreover, these organisms pose no threat to humans, which further adds to their advantage as a research object. Consequently, this cockroach can serve as a valuable resource in scientific and educational contexts, e.g. as an anatomical model [4] [5].
The body of a cockroach is also interesting from the perspective of the field known as biomimicry, particularly in terms of locomotion. This insect, despite its relatively large weight, is capable of climbing even on smooth surfaces like glass, and even upside down.
This animal has become an inspiration for me in building educational robots, such as PCHLA (eng. The Flea, Fig. 9), REKSIO, and the most advanced robot equipped with the AAM module, called MUTRA, which you can read about here.

Fig.9 - Educational walking robot
Thanks to Mr. Fruitman (G. Gruszka) for the donation of the first specimen and assistance with the documentation process.
The death's head cockroach Blaberus craniifer is a cockroach species that belongs to the Blaberidae family. It is frequently mistaken for the discoid cockroach Blaberus discoidalis, due to its similar look. It can be identified by its wings adorned with a jet black cloak-like pattern and a skull-shaped mark on its pronotum (Fig.10). The name "death's head" originates from this structure (lat. cranii, "of the head," and lat. fer, which means "carry"). Because of their distinct appearance and specific characteristics, these cockroaches are popular as low-maintenance pets or display insects among entomologists and hobbyists like me.

Fig.10 - Female Blaberus craniifer
With an average body length of 5.0 to 5.3 cm (2.0 to 2.1 in), and a total length of 6 to 6.5 cm (2.4 to 2.6 in), including the wings, this species belongs to the larger cockroaches. In exceptional cases, females can reach a body length of up to 6.2 cm (2.4 in). The males have a slightly narrower body, measuring approximately 1.8 cm (0.7 in) in width compared to the females' average width of 2.1 cm (0.8 in). Both genders have antennae measuring about 3 cm (1.2 in) in length.
Juvenile individuals are more oval-shaped and wingless (Fig.11).

Fig.11 - Juvenile Blaberus craniifer
This insect naturally occurs in Mexico, the West Indies, and Central America. Additionally, it has been introduced to southern Florida in the United States. These cockroaches can be found on the forest floors, where they hide among leaf litter and decaying wood. When faced with a predator's presence or an actual attack, this cockroach employs a defensive strategy of burrowing with its head and pronotum into softer surfaces whenever feasible.
Adults of Blaberus craniifer, unlike completely wingless Madagascar hissing cockroaches, possess wings with pink flight muscles that enable them to engage in sustained flight. Nevertheless, their typical behavior involves gliding over short distances rather than extended flights (Fig.12). On the other hand, juvenile individuals do not have wings and display a preference for burrowing instead.
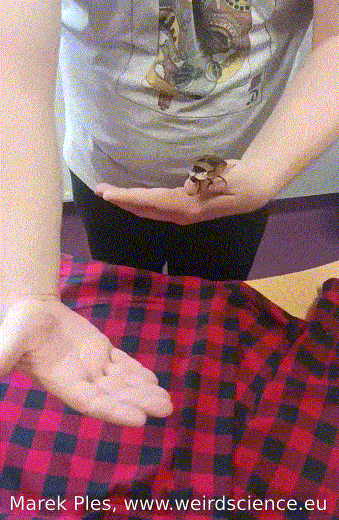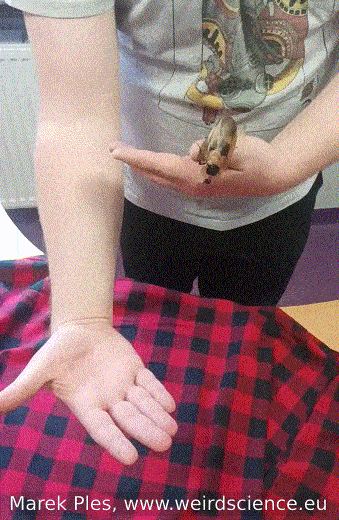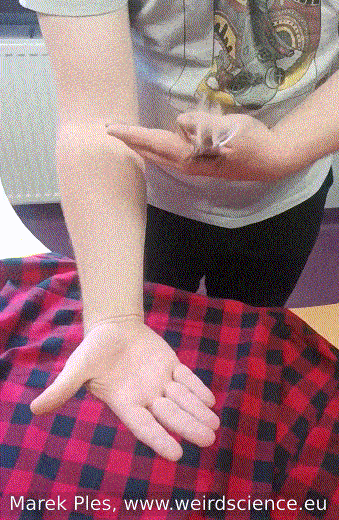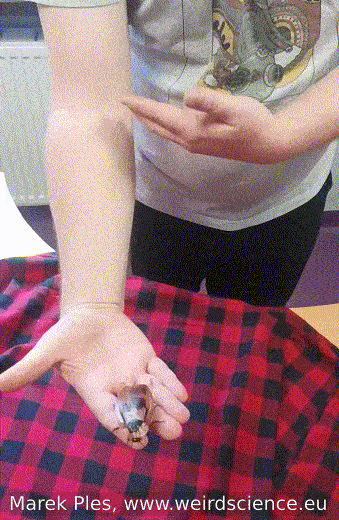
Fig.12 - Flight of the cockroach from hand to hand
This species of cockroach, as similar to the one mentioned in the previous paragraph, has proven to be highly valuable in scientific research (including neurobiology) and education [6] [7].
Porcellio scaber is a woodlouse species that is native to Europe but has a global distribution. It is commonly encountered in significant numbers across various regions, and many predators, such as shrews, centipedes, toads, spiders, and even some birds, prey upon them. Porcellio scaber can be found throughout Central and Western Europe. In the United Kingdom, it is one of the "big five" woodlouse species, which includes the common pill woodlouse Armadillidium vulgare, common shining woodlouse Oniscus asellus, common striped woodlouse Philoscia muscorum and common pygmy woodlouse Trichoniscus pusillus [8]. It has also successfully colonized regions like North America, South Africa, and even remote areas such as the sub-Antarctic Marion Island, largely due to human activities. Additionally, Porcellio scaber is the most widespread woodlouse species found in Australia.
The common rough woodlouse has an oval-shaped body that can reach a length of up to 20 mm (0.79 in). It typically exhibits a gray color, with a paler shade on its underside. However, it's also possible to observe variations in color, such as brown, blue, yellow, orange, or even a hint of pink. The upper surface of its segmented exoskeleton is covered in small bumps called tubercles, which contribute to its common name (Fig.13). Moving to its head, Porcellio scaber possesses two pairs of antennae, with the inner pair being quite small. On the dorsal side of the head, you can find two compound eyes, while the mouthparts are located on the ventral side. It has a total of seven pairs of legs, corresponding to the seven segments of its thorax. The abdomen is relatively short and comprises six segments. On the ventral side of the abdomen, there are two whitish structures known as pseudo-lungs, which are connected to the outside air through pores. At the rear end, there is a small structure called a telson, flanked by a pair of appendages referred to as uropods.

Fig.13 - The common rough woodlouse
It inhabits similar places to the common woodlouse Oniscus asellus. It is most common in gardens, at the edges of meadows, and in sunny forests (Fig.14).

Fig.14 - The common rough woodlouse is going for a walk
During the day, it hides under stones and decaying wood (Fig.15).

Fig.15 - Under stone
Porcellio scaber respond to certain stimuli with kinesis behavior. To attempt to find an environment where they lose less moisture and then stay there, animal alter factors such as speed (orthokinesis) and rate of turning (klinokinesis). When in a dry or hot environment, these woodlice have been observed increasing speed and turn more often in an attempt to leave the unfavorable environment. In a moist, dark, cool environment, they slow down dramatically and often stop altogether. This can be proven in a simple way by placing several woodlice in a chamber, where one half is lined with moist cotton and the other half with a desiccant (Fig.16A). Then, perpendicular to the boundary between the moist and dry environments, we must cover half of the chamber using black paper (Fig.16B).

Fig.16 - Woodlice Choice Chamber Experiment
In this way, we divide the chamber into four zones:
As can be seen (Fig.17), the crustaceans of our interest tend to gather most frequently in zone II, less commonly in zone III, and zones I and IV are only sporadically chosen by them. It is clearly evident that these organisms prefer moist habitats and, whenever possible, moderately or even low illuminated ones [9].
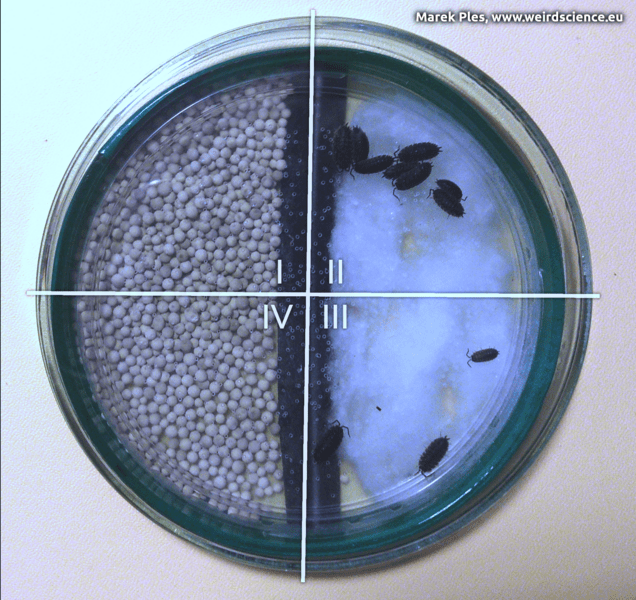
Fig.17 - Effect of experiment
In addition, Porcellio scaber exhibits a reflex called turn alternation, where it alternates between making left and right turns during klinokinesis. This behavior plays a crucial role in facilitating effective escape from predators. The alternating turns contribute to forming a straight line, leading to more efficient movement away from the threat. In contrast, random turns have the potential to result in circular paths that could bring the woodlouse back towards the predator [10]. It is easy to observe if we place this woodlouse into a simple maze; at each intersection, it indeed turns in the opposite direction from the previous one (Fig.18).

Fig.18 - Porcellio scaber in the simple maze
The common rough woodlouse is a beautiful and, at the same time, highly interesting and easy-to-keep organism. It can be useful in research within the fields of neurobiology, behavioral science, and other disciplines.
Daphnia pulex, widely distributed across the Americas, Europe, and Australia, holds the title of being the most common water flea species (Fig.19). This species, recognized as a model organism, achieved a significant milestone by being the first crustacean to have its genome fully sequenced.

Fig.19 - Daphnia pulex
Daphnia pulex is an arthropod with indistinguishable body segments. Identification is possible through appendages and internal anatomy. Its head has six fused segments with mouthparts and two pairs of antennae. It has five pairs of appendages with no clear division between the thorax and abdomen. The shell extends into a spine (Fig.20). This tiny crustacean reproduces through cyclical parthenogenesis, alternating between sexual and asexual reproduction [11].
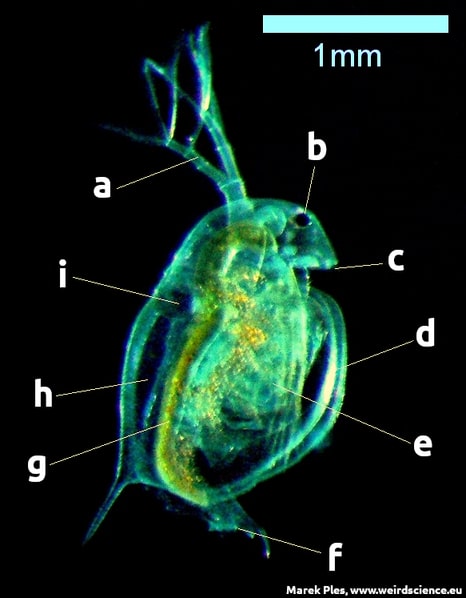
Fig.20 - Anatomy of the Daphnia pulex; a - second pair of antennae, b - compound eye, c - first pair of antennae, d - carapace, e - thoracic legs, f - anus, g - digestive tract, h - brood chamber, i - heart; dark field
Daphnias have a well-developed second pair of antennae. As you can see, they are very large and highly branched. What is their purpose? Well, these antennae serve as the primary organ of movement for this orgamism. They use them to perform paddling movements, allowing these crustaceans to move in the water in the jump-like manner - that's why they are commonly called water fleas.

Fig.21 - Daphnia pulex individuals jumping in water
Daphnia have an open blood circulation. The heart is located dorsally. Blood cells - just like the entire heart and other structures - are easily visible through the transparent body as they flow rapidly through the body cavity (Fig.22). At 20°C, it beats about 200 times per minute, slowing down at lower temperatures [12].

Fig.22 - Beating heart of Daphnia
Water fleas are easy to breed, harmless to humans, and their bodies are transparent, making them an excellent experimental subject.
The antlions comprise approximately 2,000 insect species belonging to the neuropteran family Myrmeleontidae. Their larvae exhibit predatory behavior by constructing pits in order to trap ants or other prey. In North America, these larvae are occasionally called doodlebugs, owing to the marks they create in the sand [13]. The adult antlions, are not as well recognized due to their shorter lifespans in comparison to the larvae. Typically active during dusk or nighttime, these adults might be misidentified as dragonflies.
Antlions can be found worldwide, with the tropics hosting the greatest species diversity. Nonetheless, a handful of species also inhabit cold-temperate regions, including the European Euroleon nostras, which I would like to tell about. Their preferred habitats are typically dry and sandy, as these conditions make it easier for the antlion larvae to dig their characteristic pits.
The antlion larva possesses a sturdy, spindle-shaped body with a somewhat plump abdomen, accompanied by a thorax that carries three pairs of walking legs. The prothorax forms a slender and mobile base for the large, square, flattened head. This head features an enormous pair of sickle-shaped jaws with sharp, hollow projections. The jaws are comprised of the maxillae and mandibles. Each mandible contains a deep groove where the maxilla fits snugly, creating a closed channel for injecting venom to immobilize the prey and enzymes to digest its soft tissues. The larva is covered in bristles that point forward, aiding in anchoring itself and providing better traction. This adaptation enables the larva to overpower prey significantly larger than itself (Fig.23).

Fig.23 - Larva of Euroleon nostras; top - dorsal view, bottom - ventral view
What may seem a bit strange, antlion larvae differ from other insects as they lack an anus. Instead, all the metabolic waste produced during the larval stage is stored in the body of insect. Some of it is utilized to spin the silk for the cocoon, while the remainder is eventually expelled as meconium at the end of the pupal stage.
The larva of Euroleon nostras creates a pit approximately 5 cm (2 in) deep and 7.5 cm (3 in) wide by digging backwards with its abdomen acting as a shovel. It marks the chosen location with a circular groove and uses one front leg to pile up loosened soil on its head, tossing each pile aside with a quick movement. This process continues as it moves gradually from the edge to the center, deepening the pit. The pit is lined only with fine grains when the slope angle reaches the critical angle of repose. By constructing the pit in a spiral, the antlion reduces the time required to complete it. Once the pit is finished, the larva settles at the bottom, buried in the soil with only its jaws exposed, often in an open position on either side of the very tip of the cone (Vid.2) [14].
Vid.2 - Antlion larvae behavior
Antlion larvae are not territorial - each occupies only as much space as is necessary to create their trap. As a result, they often occur in large aggregations.

Fig.24 - Multiple traps in a small area
The antlion's steep-sloped trap, designed to guide prey into its mouth while avoiding sand avalanches, is a remarkably simple and and one of the most efficient mechanism found in the animal kingdom. The lining of fine grains ensures that prey-carrying avalanches are as large as possible. The loose sand on the pit's sides provides an insecure foothold for unsuspecting insects like ants that venture over the edge. Once the prey slips to the bottom, the lurking antlion quickly seizes it. If the prey attempts to climb the walls of the pit, it is promptly hindered and brought down by showers of loose sand thrown by the larva from below. Additionally, by undermining the pit's sides with loose sand from the bottom, the larva causes them to collapse, taking the prey along with them.
Vid.3 - Antlion and ant
Antlion larvae possess the ability to capture and kill various insects and other arthropods, including small spiders. The hollow projections in their jaws serve to extract liquidified insides from their prey. Once the contents are consumed, the dry remains are discarded from the pit (Fig.25). The larva then prepares the pit anew by removing collapsed material from the center.

Fig.25 - Euroleon nostras discards the remains of an ant from the trap
Once the larva reaches its maximum size, it enters the pupal stage and undergoes metamorphosis. It creates a spherical cocoon using sand or local substrate held together with fine silk produced by a slender spinneret located at the rear of its body. The cocoon is often buried several centimeters deep in the sand. After approximately one month, the larva completes its transformation into an adult insect. It emerges from the cocoon and within around twenty minutes the adult's wings fully extend, enabling it to take flight in search of a mate [15]. Adult antlions are significantly larger than their larval stage, exhibiting one of the largest size differences between this development stages among all holometabolous insects. This disparity is due to the adult's exoskeleton being remarkably thin, fragile, and exceptionally lightweight (Fig.26).

Fig.26 - Euroleon nostras, imago
I must admit honestly that antlions belong to the group of my favorite representatives of the class Insecta.
For more information, please visit www.weirdscience.eu. You can also contact me by email (moze.dzis@gmail.com).