Skrzyp - roślina z przeszłości
Poniższy artykuł został opublikowany pierwotnie w czasopiśmie dla nauczycieli Biologia w Szkole (4/2016):

Paleontologia jest dziedziną wiedzy zajmującą się organizmami kopalnymi. Dzięki niej możemy wnioskować na podstawie badania skamieniałości i innych śladów pozostawionych przez organizmy żyjące w przeszłości o tym, jak wyglądało życie na Ziemi w minionych epokach geologicznych.
Interesujące jest, że kiedy mówimy o skamieniałościach, to mamy na myśli najczęściej jedynie pozostałości zwierzęce. A przecież wśród organizmów kopalnych możemy znaleźć także bardzo interesujące grupy roślin. Niektóre z nich wymarły całkowicie, np. ryniofity Rhyniophyta, będące prawdopodobnie pierwszymi roślinami lądowymi, wywodzące się z nich trymerofity Trimerophyta, późniejsze zosterofilofity Zosterophyllophyta i wiele innych [1].
Istnieją też grupy roślin, które mimo że dawniej były bardzo liczne, to dziś są reprezentowane przez niewielką liczbę gatunków. Tak jest w przypadku skrzypowych Equisetopsida - większa część znanych gatunków należących do tej klasy to rośliny kopalne. Należy tu wymienić praskrzypy Hyeniales, klinolisty Sphenophyllales, czy kalamity Calamitales [2]. Te ostatnie były okazałymi roślinami o pokroju drzewiastym. Ich wysokość dochodziła do 30m i były ważnym elementem lasów w karbonie (około 300mln lat temu).
Wszystkie żyjące obecnie skrzypowe należą do rodzaju skrzypów Equisetum obejmującego jedynie 15 gatunków [3]. Jako gatunek typowy przyjęto skrzyp bagienny Equisetum fluviatile. W środowisku naturalnym Polski spotkać można także innych przedstawicieli tego rodzaju, m.in. skrzyp olbrzymi Equisetum telmateia, skrzyp leśny Equisetum sylvaticum, skrzyp zimowy Equisetum hyemale, skrzyp łąkowy Equisetum pratense i inne.
Skrzypy z wielu powodów są bardzo interesującymi roślinami. Dlatego myślę, że zasługują one na to by przyjrzeć się im bliżej.
Obserwacje
W niniejszym artykule opiszę obserwacje, jakie można poczynić w odniesieniu do często występującego skrzypu polnego Equisetum arvense. Oczywiście nic nie stoi na przeszkodzie, by badać także inne gatunki, ale wybrałem właśnie ten, ponieważ można go spotkać w łatwo dostępnych miejscach, takich jak np. wilgotne łąki, skarpy, nasypy kolejowe i inne.
Skrzypy wytwarzają kłącza, które rosną mniej więcej poziomo na pewnej głębokości pod ziemią. Z kłączy wyrastają pionowo nadziemne pędy. Pędy pod- i nadziemne są tworzone przez występujące na przemian człony długie (międzywęźla) i krótkie (węzły). Z węzłów kłącza wyrastają korzenie, natomiast z węzłów pędu nadziemnego liście i rozgałęzienia pędu. Budowa pędu skrzypu jest charakterystyczna: położona w centrum luźna tkanka miękiszowa w miarę wzrostu ulega degeneracji, co pociąga za sobą powstanie dużego kanału (Fot.1). Jak widać, wokół niego rozmieszczone są też mniejsze kanały: położone w obrębie kory pierwotnej kanały walekularne, natomiast w sąsiedztwie wiązek przewodzących kanały karynalne [4].

Specyficzne dla tej grupy roślin jest to, że zarówno liście, jak i odgałęzienia są osadzone zawsze w okółkach.
U wielu przedstawicieli rodzaju skrzypów, m.in. u opisywanego tutaj skrzypu polnego, każde kłącze wytwarza w ciągu roku dwa rodzaje pędów nadziemnych: zarodnionośne i płonne (asymilacyjne).
Pędy zarodnionośne pojawiają się wiosną, najczęściej w okresie od marca do maja i osiągają wysokość kilkunastu centymetrów. Ich wygląd pozwala natychmiast uwierzyć, że skrzypy są pozostałością świata roślin minionych epok. Łodygi roślin są bezzieleniowe, praktycznie pozbawione chlorofilu – korzystają z substancji odżywczych zgromadzonych w kłączu (Fot.2).

Drobne liście wyrastające z węzłów są zrośnięte w kielichowate pochwy, czasem lekko rozdęte. Najbardziej rzucającym się w oczy szczegółem budowy pędu jest osadzony na jego szczycie kłos zarodnionośny nazywany inaczej strobilem (Fot.3).

Na przekroju (Fot.4) można zauważyć, że kłos zbudowany jest z tarczowatych elementów osadzonych na trzoneczkach. Są to specyficznie ukształtowane liście zarodnionośne [5].

Przy bliższym przyjrzeniu się (Fot.5) można zauważyć, że pod każdym tarczowatym liściem znajdują się przyrośnięte do niego zarodnie. Są one otoczone jednowarstwową osłoną otwierającą się podłużną szczeliną w stronę trzonka liścia zarodnionośnego. W zarodniach powstają zarodniki.

Pędy zarodnionośne skrzypu polnego żyją najczęściej bardzo krótko, czasem jedynie kilkanaście dni. Obumierają po dojrzeniu zarodni i wysypaniu zarodników.
Po obumarciu pędów zarodnionośnych rozwijają się silnie rozgałęzione pędy płonne o wysokości do 50cm (Fot.6).

Pędy płonne są zielone. Posiadają więc chlorofil i prowadzą fotosyntezę. Głównymi organami asymilacyjnymi nie są słabo rozwinięte i niewielkie liście zrośnięte w pochwy otaczające węzły pędu (Fot.7), lecz zielona łodyga i pędy boczne.

Skrzyp gromadzi składniki odżywcze w podziemnym kłączu aż do jesieni, kiedy pęd nadziemny obumiera.
Niektóre skrzypy, np. skrzyp leśny, wytwarzają bezzieleniowe pędy zarodnionośne ze strobilem, które po wytworzeniu zarodników nie obumierają lecz rozgałęziają się, a jednocześnie zaczynają produkować chlorofil i prowadzić fotosyntezę. Natomiast skrzyp zimowy wykształca tylko jeden rodzaj pędów. Są one nierozgałęzione, zielone i wytwarzają kłos zarodnionośny na szczycie.
Zarodniki skrzypu także wykazują pewne ciekawe własności. Są kuliste, jednakowe, o średnicy około 50μm. Rolą zarodników jest rozmnażanie bezpłciowe. Rozwijają się nich gametofity męskie (zawierające plemnie) i żeńskie (wykształcające rodnie). Przedrośla skrzypów są więc dwupienne. Gametofity są niewielkie, a ich wymiary nie przekraczają kilku milimetrów. Do zapłodnienia dochodzi za pośrednictwem wody, po czym rozwija się sporofit wykształcający opisane uprzednio pędy płodne i płonne [6].
Pozyskanie materiału do doświadczeń nie jest trudne – wystarczy zebrać pędy zarodnionośne skrzypu. Po podsuszeniu i lekkim potrząśnięciu wysypuje się z nich duża ilość zarodników w postaci szarozielonego puszystego proszku (Fot.8).

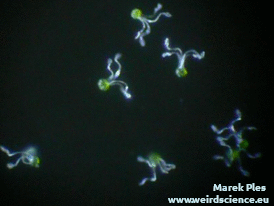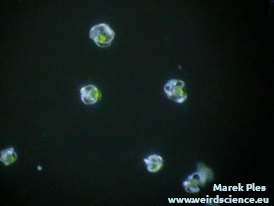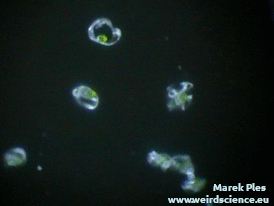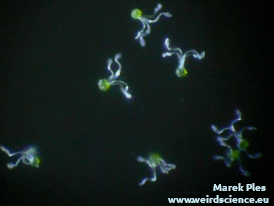
Każdy zarodnik jest wyposażony w cztery wstęgowate wyrostki. Są to tak zwane elatery, czasem nazywane też hapterami (sprężycami) [7]. Elatery wykazują bardzo wyraźne ruchy higroskopowe. W suchym powietrzu prostują się (Fot.9A), natomiast w wilgotnym skręcają ciasno wokół zarodnika (Fot.9B).

Zmiany kształtu elater mogą być tłumaczone ich dwuwarstwową strukturą. Wewnętrzna warstwa ściany zawiera gęsty układ ułożonych wzdłużnie mikrofibryl celulozowych. W bardziej zewnętrznej warstwie układ mikrofibryl jest znacznie luźniejszy [8]. Obie warstwy reagują na obecność wody zróżnicowaną zmianą objętości, co pociąga za sobą wygięcie całego elementu.
Taki plan budowy struktur wykazujących ruchy higroskopowe jest charakterystyczny – podobny można spotkać także u innych roślin. Przykładem mogą być np. łuski okrywy kwiatostanu kocanek ogrodowych Xerochrysum bracteatum, czy łuski szyszek żeńskich świerka Picea [9].
Uważa się, że opisana cecha budowy zarodników skrzypu jest przystosowaniem do anemochorii (wiatrosiewności). Wyprostowane w suchym powietrzu elatery zwiększają wydatnie powierzchnię zarodników, czym ułatwiają ich rozprzestrzenianie przez wiatr. Dodatkowo, za ich pomocą zarodniki mogą się łączyć w większe zespoły przenoszone drogą powietrzną na duże odległości.
Ruchy higroskopowe mogą mieć w tym przypadku także jeszcze inną, dosyć zaskakującą funkcję. Okazuje się, że przy cyklicznych zmianach wilgotności powietrza zarodniki mogą wykonywać za pomocą elater ruchy typu kroczącego! Widać to doskonale na na Fot.10. Przy każdej zmianie wilgotności elatery wyginają się i prostują, co skutkuje zauważalnym przemieszczaniem się zarodników (porównaj Fot.10A i Fot.10B). Ruch ten nie wymaga wydatkowania energii, ale pozwala na pokonywanie stosunkowo dużych odległości – zarodnik „a” przebył drogę 98μm przemieszczając się na odległość 76μm, czyli wielokrotnie więcej niż wynoszą jego wymiary. Trzeba jednak zaznaczyć, że ruch ten zachodzi w przypadkowych kierunkach. Jest to szczególnie dobrze widoczne na przykładzie zarodnika „b”, który pokonał drogę 102μm, ale przemieścił się jedynie o około 10μm.

To jednak nie wszystko – przy szybkim spadku wilgotności zarodniki mogą nawet wykonywać skoki o wysokości dochodzącej do 1cm osiągając przy tym prędkość około 1m/s [10]. Dużą rolę gra przy tym elastyczność odkształcających się elater.
Jak widać, higroskopowe ruchy elater mogą pełnić bardzo znaczącą rolę w rozprzestrzenianiu się zarodników opisywanej rośliny. Zauważmy, jakie skomplikowane i ciekawe mechanizmy występują u przedstawiciela skrzypów – grupy roślin uważanej za prymitywną.
Znaczenie
Skrzypy są uznawane za uciążliwe chwasty. Mogą odrastać nawet z fragmentów kłączy, dlatego rośliny te są trudne do wyplenienia.
Tkanki skrzypu zawierają duże ilości krzemionki SiO2, saponiny, alkaloidy (np. nikotynę) oraz flawonoidy. Niewielkie bulwy wytwarzane przez kłącza wykazują dużą zawartość skrobi [11].
Skrzyp polny w większych ilościach może być trujący dla zwierząt (np. gospodarskich), ale ma też pewne znaczenie jako roślina lecznicza. Ma działanie moczopędne – jest wykorzystywany w profilaktyce kamicy nerkowej, a także przy dolegliwościach układu moczowego. Uważa się, że skrzyp może być pomocny przy powstrzymywaniu krwawień. Wykazuje też dobry wpływ na stan naczyń krwionośnych, błon śluzowych, tkanki łącznej, kości, paznokci i włosów [12] [13].
Dzięki dużej zawartości krzemionki, a co za tym idzie dobrych właściwościach ściernych, skrzyp był dawniej dosyć powszechnie wykorzystywany do czyszczenia i polerowania naczyń metalowych oraz broni.
Literatura:
- [1] Taylor T.N., Taylor E.L., Krings M., Paleobotany - The Biology and Evolution of Fossil Plants, Academic Press, 2009 powrót
- [2] Jachowicz A., Dybova-Jachowicz S., Paleobotanika, Wydawnictwo Uniwersytetu Śląskiego, 1994 powrót
- [3] Hassler M., Swale B., World species list, dostępne online: http://homepages.caverock.net.nz/~bj/fern/equisetum.htm [dostęp: 14.06.2016] powrót
- [5] Szweykowska A., Szweykowski J., Botanika, tom 2 – Systematyka, Wydawnictwo Naukowe PWN, Warszawa, 2009, str. 256 powrót
- [6] Ibid., str. 249 powrót
- [7] Podbielkowski Z., Rejment-Grochowska I., Skirgiełło A., Rośliny zarodnikowe, Państwowe Wydawnictwo Naukowe, Warszawa, 1986 powrót
- [8] Beer R., The development of the spores of Equisetum, New Phytologist, 1909, 8(7), str. 261 – 266 powrót
- [9] Uehara K., Kurita S., An ultrastructural study of spore wall morphogenesis in Equisetum arvense, American Journal of Botany, 1989, 76, str. 939 – 951 powrót
- [10] Ples M., A jednak się porusza! Ruchy higroskopowe roślin, Biologia w Szkole, 3 (2016), Forum Media Polska Sp. z o.o., str. 52 – 56 powrót
- [11] Marmottant P., Ponomarenko A., Bienaime´ D., The walk and jump of Equisetum spores, Proceedings of Royal Society, 2013, 280 powrót
- [12] Mowszowicz J., Krajowe chwasty polne i ogrodowe, Państwowe Wydawnictwo Rolnicze i Leśne, Warszawa, 1986, str. 30 – 32 powrót
- [13] Rybak D., Atlas - Rośliny lecznicze, Wydawnictwo Arkady, Warszawa, 1993, str. 249 powrót
Autorem fotografii jest Marek Ples.
Uzupełnienie autora
Jako dodatek do powyższego artykułu chciałbym zaprezentować film:
Marek Ples